

| Journal of Hematology, ISSN 1927-1212 print, 1927-1220 online, Open Access |
| Article copyright, the authors; Journal compilation copyright, J Hematol and Elmer Press Inc |
| Journal website http://www.thejh.org |
Original Article
Volume 4, Number 2, June 2015, pages 164-170
Treatment of Anemia Owing to Increased Menstrual Blood Loss: Activity of Physiological Modulators
Umberto Cornellia, b, d, Gianni Belcaroc
aLoyola University School of Medicine, Chicago, IL, USA
bPiazza Novelli, 5 20129 Milan, Italy
cIrvin Labs Spoltore, University of Chieti, Italy
dCorresponding Author: Umberto Cornelli, Department of Molecular Pharmacology and Therapeutics, Loyola University School of Medicine, Chicago, IL, USA
Manuscript accepted for publication June 29, 2015
Short title: Anemia Due to Menstrual Blood Loss
doi: http://dx.doi.org/10.14740/jh215w
| Abstract | ▴Top |
Background: Heavy menstrual bleeding can lead to iron-deficiency anemia. The objective was to ascertain whether daily administration of a formula containing iron, essential amino acids, ascorbic acid and B-group vitamins for 28 days will increase hemoglobin levels by 0.5 g/dL and reduce menstrual bleeding. The formula (F) was conceived according to the physiological modulation concept to increase Hb levels and protect Hb from oxidation.
Methods: This is a single blind, placebo-controlled clinical trial on 26 anemic women (Hb < 12 g/dL) with heavy menstruation. Hemoglobin, iron balance, cell count and menstrual bleeding were measured after treatment with placebo and F.
Results: A much higher quantity of Hb was generated by treatment with F than expected (2.6 g/dL instead of 0.5 g/dL). The iron-related variables (iron, ferritin and transferrin), cell count (erythrocytes and platelets) and erythrocyte characteristics (mean corpuscular volume (MCV) and mean corpuscular hemoglobin (MCH)) improved. Menstrual blood flow decreased significantly.
Conclusion: There is no clear explanation of the results unless the F stimulates the mobilization of a “hidden” iron reserve, not detectable by ferritin levels, and triggers Hb synthesis and a decrease in menstrual bleeding.
Keywords: Iron deficient anemia; Menstrual flow; Physiological modulators
| Introduction | ▴Top |
Heavy menstrual bleeding is a common problem among women of reproductive age, which has been reported in some national surveys [1-3] as affecting between 13% and 19% of women.
Heavy bleeding during menstruation is accompanied by other symptoms, particularly iron-deficiency anemia and pain that interferes greatly with daily life and leads to a substantial economic burden [2].
Median blood loss is estimated to be about 30 mL [4] and the 75th, 90th and 95th percentile values are 52.4, 83.9 and 118.0 mL respectively. Values > 80 mL are usually considered as heavy bleeding or menorrhagia.
Excessive bleeding causes a decrease in iron stores in the body, and a negative iron balance is slowly induced, followed by insufficient iron supply to erythrocyte precursors. The final result is a decrease in Hb concentration.
An Hb concentration < 12 g/dL is considered the lowest normal value in women. Before the Hb decrease becomes evident, it is possible to measure the iron stores through ferritin levels and values between 12 and 15 ng/mL indicate iron store deficiency.
The data in the literature [4] indicate that the physiological iron requirements of adult women consist of the replacement of the iron contained in cells lost from the exterior and interior surfaces of the body (14 μg/day/kg ± 15%) plus the iron lost through menstruation. The total daily iron requirements for the 95th percentile correspond to 2.84 mg/day. Higher values are calculated for teenagers: up to 3.21 mg/day.
These data are calculated considering a mean menstrual interval of 28 days and an Hb iron quantity of 3.34 g (our calculation of hemoglobin iron content is 3.64 g, as we will explain below).
Some authors give figures for iron loss during menstrual bleeding of between 4 and 37 mg/period [5]. The highest value is consistent with the 90th percentile of the previous author [4].
Other authors calculate the loss of iron during menstrual bleeding as 2.32 mg/day [6] on average, and one author came up with different figures a few years earlier [7].
The differences between the figures are not surprising since median menstrual iron excretion is highly skewed. One reason is that there are several single-nucleotide polymorphisms (SNPs), which may have different effects on serum transferrin levels, degree of saturation and number of transferrin receptors for a given population [8]. Some polymorphisms have been described as being the cause of iron-deficiency anemia [9].
The iron needed to balance the loss can derive from food and/or from supplements, and may have different degrees of absorption.
The iron bioavailability in normal subjects following a western diet is 15% of intake on average (ranging from 14% to 17%). Therefore the diet should contain 18 mg of iron/day [3]. Different figures have been proposed by the FAO/WHO [10], ranging between 5% and 15%, according to the diets of different countries, such as Latin America, where values between 7.5% and 13.4% have been estimated on the basis of radio-iron absorption [11]. The impact of menstrual blood flow and diet on iron deficiency in women has been determined by many authors [12-14] and usually dietary iron intake is considered insufficient to prevent anemia.
Some authors have studied iron bioavailability in women according to diet [15, 16].
Differences of about six- to eight-fold have been measured between high iron-bioavailability diets (e.g. high vitamin C, low phytate, low calcium, rich in meat) and low iron-availability diets (e.g. low vitamin C, high phytate, high calcium, no meat) for periods of 12 weeks in the same women. Differences in iron absorption between lactoovo vegetarian and non-vegetarian diets given to the same women have also been shown [7].
Iron absorption increases when the iron reserve is depleted, as in anemia. Women with the lowest iron stores, even with normal Hb concentrations, absorb 30-35% of total dietary iron, whereas women with the highest iron stores absorb only 3% from the same high-bioavailability iron diet [6].
The treatment of anemia owing to menstrual bleeding is based on iron supplementation at variable dosages, ranging from 14 mg/day [17] to 60 mg/week or 60 mg/day only during menstruation [18].
Although the WHO recommends that all women of childbearing age take 120 mg iron pills once a week as prevention [19], for some authors the rule of thumb in the treatment of subjects who already have anemia is to administer 500 - 800 mg of iron for 4 weeks. This dose increases Hb by 2 - 3 g/dL and further replenishes iron stores. Some authors recommend continuation of this regimen for several additional months [20].
Our belief is that the amount of iron commonly used to treat anemia is too high and an appropriate combination of iron/amino acids/vitamins should be used.
The aim of this trial was to treat anemia in women with heavy menstrual bleeding using the physiological modulator concepts [21]. This consists of administering a combination of natural (physiological) compounds capable of regulating biological functions (modulation), in the smallest amount needed [22, 23].
In practice, a new formula (F) was prepared using the following components: iron, the essential amino acids contained in Hb, ascorbic acid and B-group vitamins (B1, B2, B3, B6, B9 and B12). The iron dosage of essential amino acids was calculated to cover, theoretically, the synthesis of 20 g of Hb in a period of 28 days and lead to an expected 0.5 g/dL total increase in Hb. B-group vitamins were added to improve Hb synthesis and protect it from oxidation. The relevant dosages were tentatively fixed between 20% and 35% of the nutrient reference value (NRV).
| Material and Methods | ▴Top |
Quantity of essential amino acids and iron in Hb
A theoretical production of 20 g of Hb in 28 days corresponds to about 5 mL of blood with an Hb concentration of 14 g/dL. The total amount of iron contained in 20 g of Hb was calculated considering MWs of 223.4 D (4 atoms of iron) and 64,500 D for Hb. In other words, iron accounts for 0.3463% of the MW of Hb. Our reference values for 5 mL of blood were 0.71 g Hb and 2.5 mg of iron.
The essential amino acid content in 0.71 g of Hb [24, 25] is shown in Table 1.
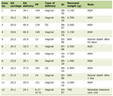 Click to view | Table 1. Quantity of Essential Amino Acids and Iron in Hb |
The total amount of iron is very close to the daily loss, according to the data reported in the literature [4, 6, 7].
Trial/subjects
Twenty-six women aged between 25 and 45 years and with 22 < BMI < 25 (kg/m2) were admitted to our single blind placebo-controlled trial. The main variables considered were the Hb concentration and the menstrual blood flow. Only women with very abundant or excessive menstrual bleeding and Hb levels between 8 and 12 g/dL were admitted (see below). Most of the women were nurse (21/26). Three menstrual periods were considered, the first and the second for enrolment and assessment of baseline variables, and the third to determine the product’s activity. During the second cycle, a placebo was given to the women, which could not be distinguished from the F (Table 2).
Click to view | Table 2. Timetable of the Single Blind Trial Conducted on Anemic Women With Very Abundant/Excessive Menstrual Flow |
Exclusion criteria
Menstrual flow with different scores during the pre-selection period was shown below. Any treatment for anemia was considered as exclusion criteria, as well as any type of gastric, neurological, or psychiatric disease and any type of cancer. Concomitant diseases such as hypertension, dyslipidemia, and diabetes type II were not considered exclusion criteria, provided the corresponding treatment had been stable for at least 2 months and did not change during the entire trial.
Product preparation
The dose of iron and essential amino acids was calculated to replace 2.5 mg of iron/day and cover the synthesis of 20 g of Hb in a period of 28 days, which was to lead to a total Hb increase of 0.5 g/dL.
The amount of iron and essential amino acids contained in a dry sachet (to be administered once day) corresponded to the quantity of Hb lost with 5 mL of blood with an Hb concentration of 14 g/dL. Ascorbic acid was included in the F in a 2:1 molar proportion to improve iron absorption [26].
The dosages of B-group vitamins were tentatively calculated between 20% and 30% of the NRV (EFSA UE document 1169/2011) in the belief that low dosages are more efficiently absorbed than high dosages [27].
According to the physiological modulators theory, the amount of each component contained in any F should be within the RDA range.
The F did not contain any calcium, magnesium or phosphate salts which may inhibit iron absorption. Furthermore, the lysine and histidine contained in the F also improve the iron bioavailability [28]. The composition is shown in Table 3.
Click to view | Table 3. Physiological Modulators Content in the Formula (F) |
Products administration
The placebo and F on trial were identical in taste and appearance (dry sachets).
The treatment with placebo (gum arabic with the same flavoring as in the product) started after the first menstrual bleeding and continued for 28 days in the form of dry sachets to be taken diluted with half a glass of water in the evening immediately before going to bed, and at least 2 h after the evening meal.
Treatment with F started immediately after the second cycle and followed the same modalities as for placebo.
The time of administrations was important and precise written instructions were given to avoid interference with the foods, and to take advantage of the peak of gastric acidity during the night which allows a better iron absorption.
Each subject was given three boxes of 10 dry sachets of each product (placebo and F) and recommendation to maintain usual diet and physical activity. There was no limitation in the use of tea or coffee. Compliance was measured by counting of the remaining sachets at the final assessment.
Sampling and variables
Fifteen milliliter blood samples were taken by venipuncture twice: after placebo (baseline) and after treatment with F. The samples were divided into three test tubes (one for serum isolation). The following variables were considered: Hb concentration (mg/dL), Ht (%) erythrocytes count (106), reticulocytes count (%), platelets count, ferritin (ng/mL), transferrin (mg/dL), mean corpuscular volume (MCV) in fL, mean corpuscular hemoglobin (MCH) in pg, and Fe serum levels (μg/dL). Venous Hb and complete blood count were analyzed using an automated instrument (Cell-Dyn 4000 Abbott Laboratories). Serum iron was determined with a chemistry analyzer (LX 20 Beckman-Coulter) as were ferritin and transferrin (Immunolite 2500, Diagnostics Product Corporation).
Menstrual blood flow
The methods used to measure menstrual bleeding were based on two variables that were measured using a score. The first variable was the menstrual bleeding which was scored from 1 to 5 based on the number of tampons and/or pads as follows: 1 = normal (< 22 tampons/pads); 2 = abundant (23 - 29 tampons/pads); 3 = high (30 - 35 tampons/pads); 5 = very high (> 35 tampons/pads) causing social/working problems.
The second variable was the duration of the menstruation which was scored from 1 to 3, where 1 means < 2 days, 2 means 3 days, and 3 means > 4 days. The total score was the sum of the two scores that could be from 2 to 8.
Statistical analysis
Hb was considered as a primary variable. Twenty is a sufficient number to obtain a power 90% with an α value of 0.05 and 1-β of 90 comparing two sets of interdependent data (Dunnett). Average values and SD were calculated for all data. The coefficient of correlations “r” between all the variables were calculated before and after the treatment taking α of 0.01 as limit of significance.
| Results | ▴Top |
All the women completed the study and there was no complaint of side effect.
The data are shown in Table 4.
Click to view | Table 4. List of the Variables Measured After 28 Days of Treatment With Placebo and After Treatment With F: average values ± SD |
The differences between the data recorded after the treatment with placebo and F are statistically significant (Dunnett) for all the variables. The cell count was improved for both erythrocytes and platelets.
Ht, MCV and MCH increased significantly but the averages were in the range of the normality even before the treatment. Reticulocytes % was very low during the placebo treatment and improved after F. The reticulocytes production index (RPI) was > 2 in one case with placebo. After F, this number increased to 6. Ferritin levels improved, indicating an increase of iron reserve for most of the women after the treatment with F.
Transferrin and serum iron levels increased of about 10% and 30% respectively. The menstrual bleeding reduction was also very consistent and was present in about 77% of the women. An inverse correlation (“r”) was found between the Hb data following placebo and the same data after F (-0.618; P < 0.01) with a more substantial effect in the women with the lowest Hb level.
No correlation was found after placebo and after F for the remaining variables with the exception of reticulocytes. The reticulocytes count before and after treatment showed a direct correlation (0.796; P < 0.01). As regards the RPI, it seems that F improves the bone marrow reaction.
| Discussion | ▴Top |
One limitation of the trial is the lack of control over iron, vitamin B and essential amino acid intake with the diet. Another limitation is the semi-quantitative measurement of the menstrual blood flow which can give only partial indications. Apart from these limitations, the results of the trial are very interesting and raise some important questions.
The subjects were asked to do not alter the quantity and type of foods, but we have no knowledge about their compliance. The only precise values were the intake of amino acids and iron needed to increase Hb by 0.5 g/dL in 28 days.
The total quantities were 70 mg of iron and 1.61 g of lysine, 1.39 g of histidine, 1.26 g of threonine, 2.42 g of valine, 0.28 g of methionine, 2.51 g of leucine, 1.08 g of phenylalanine and 0.27 g of tryptophan, respectively.
The data we observed after treatment with F were very surprising, since the improvement in Hb from 10.2 g/dL to 12.6 g/dL was much greater than expected.
The first finding to be considered is the increase in serum iron and transferrin that were obtained with a very low iron dosage. This is not surprising because iron absorption increases dramatically when iron reserves are depleted [3] as was the case of the women treated with F.
Looking at the amount of iron in relation to Hb levels rises a question: the average increase in Hb (2.4 g/dL) is about five times the expected increase (0.5 g/dL) both for the administered quantities of essential amino acids and iron.
Regarding the amino acids, the extra quantities might derive from the plasma, which contains concentrations of essential amino acids [29, 30] that are more than enough to cover needs. Even though there are no data published on the amino acids content in anemic women, their expected concentrations are more than sufficient to afford the Hb synthesis increase measured.
Furthermore, the quantity of essential amino acids that was administered is unlikely to increase the plasma level to a great degree. Again in this case the amount was (as for iron), about one-fifth of the quantity necessary to cover the Hb increase (two portions of any kind of meat can cover all the essential amino acids needed).
However, the iron data need some other considerations. The quantity of iron needed for the observed average increase in Hb is 1,903 mg, whereas the average expected level, even considering 100% absorption of the 70 mg given during the 28 days, was 1,573 mg. The difference is about 330 mg of iron; the question is “where does this high quantity of iron come from?” considering that this extra amount is equal to a theoretical complete iron reserve in a non-anemic women.
One hypothesis is that the F allows an iron reserve mobilization that cannot be fully detected with the usual methods such as ferritin. This would mean that hemosiderin contained in liver, spleen and macrophages of anemic women is “hidden” and not re-converted from hemosiderin to ferritin and then into free iron of the cytoplasmic labile pool to be used for the heme synthesis.
The continuous exchange process between hemosiderin/ferritin/labile iron pool flows in both directions and follows a density gradient [31]. The iron has to be used for the heme synthesis through a well coordinated mechanism. When Hb is needed the iron is mobilized by the stores (hemosiderin/ferritin) and shuttles between cytoplasm and mitochondria, where the final steps of heme synthesis take place [32]. After heme synthesis, it is incorporated into α and β chains [33, 34].
However, there is a fundamental step for iron utilization: it has to be solubilized because the iron in the stores (ferritin/hemosiderin) is in the form of ferrihydrite [5Fe2O3·9H2O]. In response to signals of iron deprivation, the cells move ferritin into lysosomes for autophagy. Nanocrystals of ferrihydrite are made soluble in the lysosomes thanks to the reduction by ascorbate and glutathione [5].
This means that some reducing molecules are needed to mobilize the iron kept into the cellular stores.
In erythrocyte precursors, most of the reducing capacity is used to protect Hb from oxidation. The protection mechanism belongs to the pentose phosphate cycle, which regenerates glutathione once it has lost its hydrogen to protect Hb [35]. The pentose phosphate cycle is regulated by some of the vitamin B group (B1) and allows NADH+ H+ (that needs vit B2 and B3 for its synthesis) to transform the oxidized glutathione (GSSG) into reduced glutathione (GSH).
Furthermore all the vitamin B group is necessary for the hemoglobin synthesis.
The Hb synthesis seen in this trial seems to have no explanation.
One possible hypothesis is that administering the F on an empty stomach (at least 2 h after the evening meal) means that it is not affected by the presence of the many food components (e.g. lipids, carbohydrates, fiber, proteins, amino acids and minerals). In these “clean conditions”, the presence of iron and essential amino acids in the precise proportion contained in Hb could be interpreted by enterocytes as blood loss in the gut. This condition may stimulate the enterocytes to release factors (unknown) through the brain and/or the liver which aim to increase Hb synthesis. In other words, the F acts as a “trigger” through a completely unknown mechanism.
Assuming this hypothesis is correct, what is important in the treatment of anemia is to maintain the ratio of iron to essential amino acids.
The only experiment reported in literature describing the activity of a combination of iron and amino acids to treat anemia was carried out in dogs made chronically anemic through phlebotomy [36].
Each iron/amino acid combination was studied separately. It is probable that these animals were completely iron depleted, and the iron remaining in the liver and other tissues appears to be essential to the cell life, and cannot be drawn upon to produce new Hb.
Most of the time the daily dosage of single amino acids was 1 g/day and the iron administered together with the amino acids was ranging between 5 to 40 mg/day. The ratio between amino acids and iron was from 2 to 30 times higher than in our trial. However, in few dogs treated with valine and methionine, the increase of Hb should have needed 3 - 4 times more iron than the quantity administered.
These data confirm that even in the controlled experimental anemia, the iron reserve behaves with unknown mechanisms, and amino acids administration may trigger some mechanism that improves the Hb synthesis.
In other words, the iron/amino acid ratio and kind of amino acid seem to be important in improving Hb synthesis in iron-deficiency anemia.
The other quite unexpected finding was the decrease of menstrual blood flow. Despite a semi-quantitative assessment, most of the women were reporting a reduction of the flow (about 70% of the cases) and we do not think this finding can be fortuitous.
The increase in platelets count might be one of the causes of the decrease in menstrual bleeding, although it was not possible to calculate a proper correlation because of the different types of variables (parametric for platelets and non-parametric for bleeding). The increase in erythrocytes may have some effect and ascorbic acid may improve the microcirculation. However, again in this case, the finding has no a clear explanation.
The number of reticulocytes improved following the treatment and the increase was more substantial in subjects with relatively higher percentages. One interpretation of this might be that the F was more substantial in women with relatively higher medullary reactivity. However, comparing the average reticulocytes data before and after the treatment, the differences are not very substantial (1.3 ± 0.82 vs. 1.9 ± 0.65).
The dose of vitamin B group was determined empirically between 20% and 30% of NRV. According to physiological modulation concepts, it is important to prevent daily dosages exceeding the RDA by much. It is safe to assume that the total daily quantity for most of the women was not exciding the RDA value, even with the supplements provided with the F, since overweight women (high intake of vitamins), or women suffering from gastrointestinal diseases (low intake of vitamins) were not admitted to the trial.
Conclusions
Anemic women with abundant menstrual bleeding can be treated daily with an F containing a limited amount of iron (2.5 mg), ascorbic acid, essential amino acids in the same proportion as in Hb, and some group B vitamins (B1, B2, B3, B6, B9, and B12). The doses of all the substances in the F were consistent with the concept of physiological modulation, which is based on the use of small quantities of the different substances combined together.
The data regarding the increase in Hb level and decrease of menstrual bleeding were impressive.
The improvement in Hb synthesis was much more substantial than the expected (2.4 g/dL instead of 0.5 g/dL). Menstrual bleeding also showed a substantial decrease. The results suggest that the improvement was not based on the quantities of substances administered, but is caused by an unknown mechanism that triggers the Hb synthesis and a decrease in menstrual blood flow.
The data need further confirmation and a larger number of cases treated for a longer period of time.
Competing Interest
The authors declare they have not competing interest.
| References | ▴Top |
- Cote I, Jacobs P, Cumming D. Work loss associated with increased menstrual loss in the United States. Obstet Gynecol. 2002;100(4):683-687.
doi - Tanaka E, Momoeda M, Osuga Y, Rossi B, Nomoto K, Hayakawa M, Kokubo K, et al. Burden of menstrual symptoms in Japanese women: results from a survey-based study. J Med Econ. 2013;16(11):1255-1266.
doi pubmed - Dorsey KA. Menorrhagia, active component service women, U.S. Armed Forces, 1998-2012. MSMR. 2013;20(9):20-24.
pubmed - Hallberg L, Rossander-Hulten L. Iron requirements in menstruating women. Am J Clin Nutr. 1991;54(6):1047-1058.
pubmed - Linder MC. Mobilization of stored iron in mammals: a review. Nutrients. 2013;5(10):4022-4050.
doi pubmed - Hunt JR. High-, but not low-bioavailability diets enable substantial control of women's iron absorption in relation to body iron stores, with minimal adaptation within several weeks. Am J Clin Nutr. 2003;78(6):1168-1177.
pubmed - Hunt JR, Roughead ZK. Nonheme-iron absorption, fecal ferritin excretion, and blood indexes of iron status in women consuming controlled lactoovovegetarian diets for 8 wk. Am J Clin Nutr. 1999;69(5):944-952.
pubmed - Blanco-Rojo R, Toxqui L, Lopez-Parra AM, Baeza-Richer C, Perez-Granados AM, Arroyo-Pardo E, Vaquero MP. Influence of diet, menstruation and genetic factors on iron status: a cross-sectional study in Spanish women of childbearing age. Int J Mol Sci. 2014;15(3):4077-4087.
doi pubmed - Pei SN, Ma MC, You HL, Fu HC, Kuo CY, Rau KM, Wang MC, et al. TMPRSS6 rs855791 polymorphism influences the susceptibility to iron deficiency anemia in women at reproductive age. Int J Med Sci. 2014;11(6):614-619.
doi pubmed - FAO/WHO joint expert consultation report. Requirements of vitamin A, iron, folate and vitamin B-12. FAO food and nutrition series 23. Rome: Food and Agriculture Organization, 1988.
- Acosta A, Amar M, Cornbluth-Szarfarc SC, Dillman E, Fosil M, Biachi RG, Grebe G, et al. Iron absorption from typical Latin American diets. Am J Clin Nutr. 1984;39(6):953-962.
pubmed - Heath AL, Skeaff CM, Williams S, Gibson RS. The role of blood loss and diet in the aetiology of mild iron deficiency in premenopausal adult New Zealand women. Public Health Nutr. 2001;4(2):197-206.
doi pubmed - Pynaert I, Delanghe J, Temmerman M, De Henauw S. Iron intake in relation to diet and iron status of young adult women. Ann Nutr Metab. 2007;51(2):172-181.
doi pubmed - Harvey LJ, Armah CN, Dainty JR, Foxall RJ, John Lewis D, Langford NJ, Fairweather-Tait SJ. Impact of menstrual blood loss and diet on iron deficiency among women in the UK. Br J Nutr. 2005;94(4):557-564.
doi pubmed - Collings R, Harvey LJ, Hooper L, Hurst R, Brown TJ, Ansett J, King M, et al. The absorption of iron from whole diets: a systematic review. Am J Clin Nutr. 2013;98(1):65-81.
doi pubmed - Hurrell R, Egli I. Iron bioavailability and dietary reference values. Am J Clin Nutr. 2010;91(5):1461S-1467S.
doi pubmed - Milman N, Clausen J, Byg KE. Iron status in 268 Danish women aged 18-30 years: influence of menstruation, contraceptive method, and iron supplementation. Ann Hematol. 1998;77(1-2):13-19.
doi pubmed - Bani S, Hassanpour-Siahestalkhi A, Hassanpour S, Mommad-Alizadeh-Charandabi S, Mirghafourvand M, Javadzadeh Y. Comparison of two iron supplementation methods on Hemoglobin level and Menstrual Bleeding in Tabriz students. Iran J Ped Hematol Oncol. 2014;4(1):11-16.
pubmed - Joint FAO/WHO Consultation on Human Vitamin and Mineral requirements (1998: Bangkok, Thailand 21-30 Sept 1998).
- Alleyne M, Horne MK, Miller JL. Individualized treatment for iron-deficiency anemia in adults. Am J Med. 2008;121(11):943-948.
doi pubmed - Olson JA. Benefits and liabilities of vitamin A and carotenoids. J Nutr. 1996;126(4 Suppl):1208S-1212S.
pubmed - Finco A, Belcaro G, Cesarone MR. Assessment of the activity of an oral contraceptive on the levels of oxidative stress and changes in oxidative stress after co-treatment with two different types of physiological modulators with antioxidant action. Contraception. 2011;84(4):418-422.
doi pubmed - Cornelli U, Belcaro G, Ledda A, Feragalli B. Activity of some physiological modulators in reducing the side effects of levothyroxine in patients suffering from primary hypothyroidism. Panminerva Med. 2011;53(3 Suppl 1):99-103.
pubmed - Hill RJ, Konigsberg W, Guidotti G, Craig LC. The structure of human hemoglobin. I. The separation of the alpha and beta chains and their amino acid composition. J Biol Chem. 1962;237:1549-1554.
pubmed - Guidotti G, Hill RJ, Konigsberg W. The structure of human hemoglobin. II. The separation and amino acid composition of the tryptic peptides from the alpha and beta chains. J Biol Chem. 1962;237:2184-2195.
pubmed - Teucher B, Olivares M, Cori H. Enhancers of iron absorption: ascorbic acid and other organic acids. Int J Vitam Nutr Res. 2004;74(6):403-419.
doi pubmed - Recommended Dietary Allowance, 10th Ed, National Res Council (US) Washington (DC) National Academic Press (US); 1989.
- Kroe D, Kinney TD, Kaufman N, Klavins JV. The influence of amino acids on iron absorption. Blood. 1963;21:546-552.
pubmed - Lewis AM, Waterhouse C, Jacobs LS. Whole-blood and plasma amino acid analysis: gas-liquid and cation-exchange chromatography compared. Clin Chem. 1980;26(2):271-276.
pubmed - Plasma amino acids. US National Library of Medicine - National Institute of Heath. http://nlm.nih.gov/medlineplus/ency/article/003361.htm.
- Saito H. Metabolism of Iron Stores. Nagoya J Med Sci. 2014;76(3-4):235-254.
pubmed - Richardson DR, Lane DJ, Becker EM, Huang ML, Whitnall M, Suryo Rahmanto Y, Sheftel AD, et al. Mitochondrial iron trafficking and the integration of iron metabolism between the mitochondrion and cytosol. Proc Natl Acad Sci U S A. 2010;107(24):10775-10782.
doi pubmed - Weatherall DJ. The control of haemoglobin synthesis. Proc R Soc Med. 1967;60(10):1037-1041.
pubmed - Chiabrando D, Mercurio S, Tolosano E. Heme and erythropoieis: more than a structural role. Haematologica. 2014;99(6):973-983.
doi pubmed - Stromme JH, Eldjarn L. The role of the pentose phosphate pathway in the reduction of methaemoglobin in human erythrocytes. Biochem J. 1962;84:406-410.
pubmed - Whipple GH, Robscheit-Robbins FS. Amino Acids and Hemoglobin Production in Anemia. J Exp Med. 1940;71(4):569-583.
doi pubmed
This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Hematology is published by Elmer Press Inc.